
Tilbudsguide, uge 45, 2019
And og æg!
Tilbudsguide, uge 44, 2019
Svinemørbrad og mandler!
Tilbudsguide, uge 43, 2019
Havregryn og oksefilet!
Tilbudsguide, uge 42, 2019
Olivenolie, granatæble og mango!
Vi har 65665 registrerede brugere. Nyeste registrede medlem er peinoin
Vores medlemmer har i alt skrevet 2049570 indlæg i 100421 emner
2 nye indlæg i dag



Muskelarkitektur fra det store til det små
Introduktion
Jeg sad og kiggede igennem hvad jeg havde skrevet tidligere og hvad der i øvrigt lå på bb.dk, og det slog mig lige pludselig, at der slet ikke var nogen grundlæggende beskrivelse af hvordan musklerne er opbygget. Når diskussionerne bliver tekniske og går lidt højt på forummet, er det altid en fordel at have konteksten på plads og vide hvad der er hvor, og hvordan det hænger sammen med strukturerne på mindre eller større planer. Så derfor har jeg skrevet en beskrivelse af musklen fra det helt store niveau, ned til det helt lillebitte.
Mono- vs. Biartikulær muskulatur
Vi er vant til at tænke på muskler som værende noget der sidder fast på to knogler der bøjer i ét led, men ikke alle muskler går over et led. Nogle få går slet ikke over et rigtigt led (bl.a. nogle af mavemusklerne og ansigtsmusklerne og nogle går over flere led). Disse benævnes biartikulære muskler, mens de almindelige (dem der går over et enkelt led) kaldes monoartikulære. De mest prominente eksempler er:
-
De fleste af lårbiceps hovederne (alle undtaget det korte hoved af biceps femoris)
-
Det lange hoved af triceps
-
Biceps (nogle mener endda at biceps er triartikulær fordi de også tæller leddet mellem underarmsknoglerne med, men det er ikke noget alle er enige om)
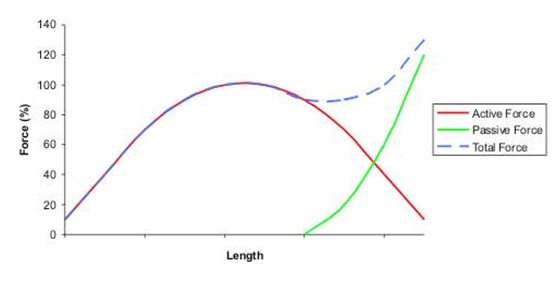
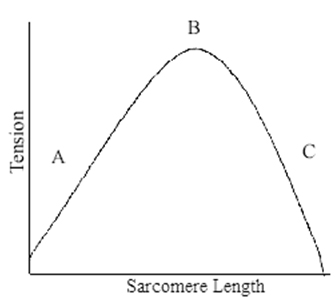
Alle muskler har en klokkeformet kraft/længde kurve. Det betyder at den kan yde mest kraft ved en bestemt længde, og mindre kraft ved mindre eller større længder. Samtidig har bindevævene i musklen en passiv komponent, som yder en eksponentielt stigende kraft, fra en given længde og opefter.
Sammen med artikulationsforholdene, har det en konsekvens for hvilke muskler der belastes mest, i forskellige øvelser, idet centralnervesystemet programmerer vores reruktteringsstrategier så vi bruger musklerne på den mest hensigtsmæssige måde. F.eks er der stor forskel på om man bruger ballerne eller baglårene til hofteekstension, afhængig af om knæene er strakt eller bøjede. Strakte knæ = baglår, bøjede knæ = baller.
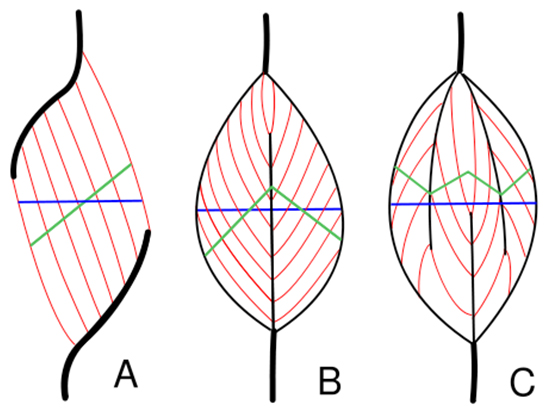
Pennat vs. ikke pennat muskulatur
Mange tænker på muskelfibre som gående igennem muskler langs med længderetningen, og jeg gik selv i lang tid, før jeg kom ind i academia, og undrede mig over de her tværgående striber man kunne se i bodybuilderes muskler.
Forklaringen er at ikke alle muskler er arrangeret med længdegående muskelfibre. Meget tydelige eksempler ser man i ekstensor muskulaturen i mennesker, altså i quadriceps, gastrocnemius og triceps.
Bundter af muskelfibre, kaldet fascikler (mere om dem senere) er simpelthen skråtstillet i forhold til den enkelte muskelstrækretning i disse muskler. Det overordnede formål med at have sådanne pennate muskler er at geare kraft/længde forholdet, idet pennation generelt giver en lidt smallere, men højere klokkekurve. På almindeligt dansk giver de altså mulighed for at producere større kraft. Man kan da også se at næsten alle mennesker er stærkere i deres ekstensor- (strækker) end i deres fleksor (bøjer) muskulatur, hvor de fleste ekstensor muskler er pennate.
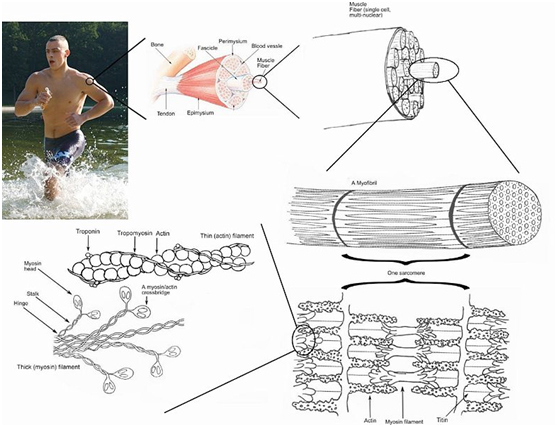
Arkitektur fra makro til mikro
Mange af kroppens væv er hierarkisk opbygget, idet at et mønster som man kan genkende på et niveau, i et eller andet omfang gentages på mindre niveauer. Det ser man også i muskler der er indkapslet i en sej bindevævshinde kaldet fascien eller epimysiet og underinddelt i fascikler.
Antallet af fascikler, på en hel muskel, svinger fra meget få og op til nogle hundrede. Hver enkelt fascikel er indkapslet i et bindevævslag kaldet perimysiet og selve fasciklen består af nogle hundrede til tusinde muskelfibre. Hver af disse muskelfibre er ligeledes indkapslet i perimysiet, og imellem dem er vævet fyldt op med noget andet bindevæv der hedder endomysiet.
Fortsætter man ind i muskelcellen (fiberen), er den fyldt op med bundter af myofibriller, som i praksis er de enheder i musklerne der får dem til at trække sig sammen. Kigger man på muskelfibre i tværsnit gennem et mikroskop vil man se, at i hvert fald mindst 95% af tværsnittet er dækket af myofibriller. For alle praktiske formål, er det altså de kontraktile proteiner der ”fylder” næsten hele musklen. I længderetningen består myofibrillerne af serielt koblede enheder der hedder sarcomerer, som jeg vil vende tilbage til senere.
I tværsnit består hver myofibril af en masse filamenter, hvor de to vigtigste er actin og myosin. Selve bevægelsen i musklerne opstår ved at myosin ”kravler” hen af actin molekylerne på bekostning af forbrug af ATP.
Det skulle være tydeligt hvad det er jeg mener, når jeg snakker om hierarki og gentagede mønstre i strukturerne på flere niveauer.
Cellerne i musklen - Muskelfibre og satellitceller
Muskelfibrene kan inddeles i hurtige (type II) og langsomme (type I). Heraf kan de hurtige yderligere inddeles i IIA og IIX, hvor IIX er lidt stærkere, noget hurtigere og meget mindre udholdende end IIA. Der findes også en tredje, endnu hurtigere type, IIB, som man dog ikke rigtigt ved om mennesker udtrykker i nogle væv. Konsensus er indtil videre at det gør vi ikke. Den findes dog i gnavere og andre mindre dyr, da kontinuummet af muskelfibre generelt er forskudt mod de hurtigere i mindre dyr, i forhold til mennesker.
Generelt ændres fibertyper ikke mellem hurtige og langsomme indenfor normale fysiologiske stimuli. Ved kronisk (uger til måneder) elektrisk stimulation kan man forårsage en overgang fra hurtige til langsomme fibre og ved total inaktivering (som ved lammelse), kan man se en overgang fra langsomme til hurtige. Det er dog sandsynligt, at hvis folk der havde været udsat for den slags stimuli, vendte tilbage til almindelige aktiviteter, så ville deres fibertypefordeling vende tilbage til deres ”normale”.
Præcis hvordan det bestemmes om en fiber skal blive hurtig eller langsom er endnu ikke særlig velkendt. Nogle af de eksisterende data antyder at en del af den population af fibre man anlægger i fostertilstanden er forudbestemt til at blive langsomme, mens en anden population kan blive til hvad som helst (både langsomme og hurtige). Derudover er er det også ud til at interaktionen med motorneuroner betyder noget, men det er stadig uklart om fibrene selv ”kalder” på de rigtige motorneuroner, eller om motorneuronerne er med til at bestemme om fibrene skal være hurtige eller langsomme, efter de har koblet sig på fiberen.
Man har en pulje af muskelstamceller i musklerne, som normalt kaldes sattelitceller. De ligger uden om muskelfibrene, men under bindevævslagene og ligger således i berøring med muskelfibrene. Når der sker skade på musklerne, aktiveres satellitcellerne, så de begynder at dele sig og nogle af dem vil fusionere med musklerne, for at gå ind og erstatte skadede cellekerner og måske opbygge fibre, der er blevet helt ødelagt.
Desværre er satellitcellers opførsel primært blevet undersøgt i muskelskademodeller, altså hvor utrænede mennesker eller dyr har været udsat for meget voldsomt arbejde og man har efterfølgende kigget på hvad der skete. Det er til gengæld svært at sige hvad deres rolle er under mere normale fysiologiske omstændigheder (hvor reel muskelskade jo er sjældent), men man ved dog at de er nødvendige for hypertrofi, og tilsvarende at en af grundene til at musklerne bliver mindre og mere tarvelige til at regenerere med alderen, er fordi satellitcellerne bliver dårligere til at gøre deres arbejde.
Under hypertrofi i forbindelse med vægttræning, menes det at i den første fase af tilpasningen sker hypertrofien primært ved at de eksisterende cellekerner programmerer syntesen af nye muskelproteiner, men kun op til en vis grænse. Det ser nemlig ud til at for at opnå hypertrofi over en vis grænse (20-30%), begynder det at blive nødvendigt at indbygge satellitceller (som bliver til nye cellekerner). Dette har foranlediget det der kaldes ”myonuclear domain theory”, hvor hver enkelt cellekerne inde i muskelfiberen kun kan vedligeholde et vist kvantum af muskelfiberen, og hvis muskelfiberen vokser udover det, er det nødvendigt med flere cellekerner.
Det er dog fortsat den dominerende teori at der ikke i et væsentligt omfang dannes flere muskelfibre i mennesker, hverken med styrketræning eller administration af anabolske stoffer, det være sig androgener, IGF, væksthormon eller andre gode sager.
Sarcomerer, costamerer og kraftoverførsel
De enkelte myofibriller i musklerne består af serielt koblede enheder af sarcomerer. Sarcomeren er fundamentalt set den mindste funktionelle kontraktile enhed i musklerne. I hver ende af sarcomer er der et Z-disk hvori actinen er forankret. I midten af sarcomeren er der en tilsvarende disk som ikke har noget selvstændigt navn, men som ligger i et område, kaldet H zonen. I denne disk er der forankret myosin, som stritter ud til hver sin side og ligger sig imellem actin filamenterne. Der findes selvfølgelig hundredevis af andre proteiner som hjælper med at holde styr på denne struktur, men nu holder vi bare fast i actin og myosin, som de vigtigste.
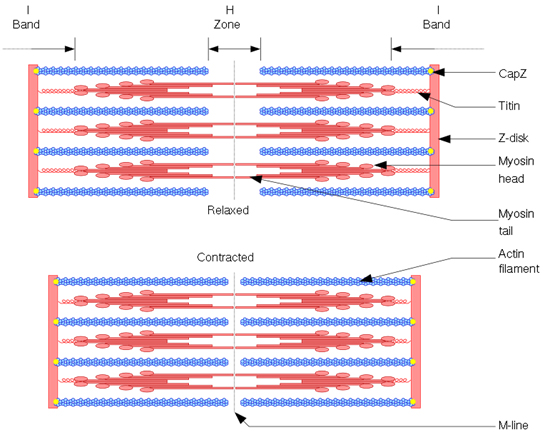
På grund af denne overlappende struktur har man målt kraft/længde forhold for costamerer selvstændigt, og her kan man observere det hierarki jeg talte om tidligere vedrørende musklernes struktur igen, for sarcomerernes kraft/længde forhold er også klokkeformet. Dette kan forklares ved at når der er 100% overlap imellem actin og myosin, er der det største potentiale til at generere kraft, men når der er mindre overlap, enten på grund af de er for langt inde over hinanden eller for langt fra hinanden, så er kraften mindre.
I myofibriller der ligger ved siden af hinanden, kan man se at Z diskene ligger lige ud for hinanden, hvilket gør at muskelfibre overordnet set forekommer stribede når man ser dem i mikroskop (heraf navnet tværstribet muskulatur, som mange også bruger). Grunden til at de er arrangeret således er at Z-diskene imellem myofibrillerne faktisk er bundet til hinanden. Og i de myofibriller, som ligger op ad muskelfiberens ydre cellevæg, er Z-diskene faktisk forbundet til cellevæggen og helt ud til bindevævslagene udenfor cellerne, igennem nogle proteinkomplekser kaldet costamerer. Man har faktisk fundet ud af at en stor del af kraften, måske størstedelen, som sarcomererne genererer, bliver projiceret ud til bindevævet omkring muskelfibrene og derefter langs med musklen frem for ned gennem de enkelte myofibriller og ud gennem enden af musklen.
Innervering og rekruttering
Hver enkelt muskelfiber bliver aktiveret af en nervecelle (motorneuronet), som har kontakt til flere andre muskelfibre. Motorneuronet er også en enorm celle, hvis krop ligger helt oppe i rygmarven og rækker helt ud til arme eller ben.
Denne gruppe af fibre udgør, sammen med motorneuronet, en motorenhed som enten kan være hurtig eller langsom. De langsomme motorenheder består af langsomme fibre og aktiveres på en bestemt måde, mens de hurtige enheder består af hurtige fibre (og typisk mange flere af dem) og aktiveres ligeledes på en karakteristisk måde.
Teorien for hvordan motorenhederne rekrutteres under stigende belastning kaldes ”Henneman’s Size Principle”. Selvom teorien har vist sig ikke at være gældende i en række specialtilfælde og at forklaringsmodellen Henneman brugte, sandsynligvis er forkert, så er selve den deskriptive del af modellen stadig god til at forklare hvordan de forskellige motorenheder inddrages i arbejde.
Modellen siger at ved helt lave belastninger, rekrutteres de små, langsomme motorenheder. Dette giver god mening, da de langsomme muskelfibre har meget stor træthedstolerance og fordi langt de fleste gange en muskel tændes, skal den kun yde max 5% af sin max kraft, idet det kun er små stabilitetsopgaver den skal klare. Den type aktivitet laver musklerne faktisk hele tiden. De hurtige muskelfibre ville slet ikke kunne holde til at være tændt på den måde, da de har en meget lavere træthedstolerance. Man kan også forestille sig at det ville være uhensigtsmæssigt at bruge så store motorenheder til at stabilere og korrigere, idet deres kraft måske simpelthen ville være for stor til at det var rigtigt praktisk at bruge til den slags opgaver. Efterhånden som belastningen stiger, rekrutteres flere og flere af de langsomme motorenheder, indtil de alle er tændt. Herefter begynder kroppen at rekruttere de hurtige. Ved cirka 80% intensitet er alle motorenheder i spil og yderligere kraft opnås ved det der kaldes ”rate coding”, altså hvor de individuelle pulstog fra motorneuronet til muskelfibrene kommer hurtigere efter hinanden.
Et af de tilfælde hvor Henneman’s princip bryder sammen, er under muskeludmattelse. Når muskelfibre udmattes, stiger deres aktiveringstærskel så de bliver sværere at tænde. Det betyder at under muskeludmattelse vil alle fibrene ikke længere være tændt, men i stedet så begynder muskelfibrene at rotere hvilke der er tændt og hvilke der er slukkede. Da de hurtige fibre udmattes lettere, stiger deres aktiveringstærskel mere og derfor er det sandsynligt, at i den rotation der finder sted under muskeludmattelse, vil de langsomme motorenheder tage en større del af slæbet.




