Tilbudsguide, uge 45, 2019
And og æg!
Tilbudsguide, uge 44, 2019
Svinemørbrad og mandler!
Tilbudsguide, uge 43, 2019
Havregryn og oksefilet!
Tilbudsguide, uge 42, 2019
Olivenolie, granatæble og mango!
Vi har 65712 registrerede brugere. Nyeste registrede medlem er peinoin
Vores medlemmer har i alt skrevet 2049579 indlæg i 100422 emner
1 nye indlæg i dag



The influence of eccentric strength training in the enhancement of muscle strength, explosive force (RDF) and muscular power
Introduction
Maximal muscle strength and power can be maximized by means of resistance training, however different types of training may lead to differential gains in one or the other parameter. Heavy-resistance strength training (HRST) leads to increased levels of contractile muscle strength during isometric, concentric and eccentric muscle actions of maximal voluntary effort. Corresponding effects of HRST on maximal muscle power production have been reported in the literature, while more specialized types of resistance training (i.e. plyometric exercises) appear to also induce gains in maximal muscle power production.
This brief review article presents and discus the effect of various resistance training modalities, including supramaximal eccentric training and plyometric training, on the maximal force and power production of human skeletal muscle in untrained subjects, athletes and aging individuals.
Heavy-resistance strength training
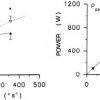
Maximal muscle strength typically is increased by 20-40% in response to HRST of medium to moderate time length (8-16 weeks) [3,13,21]. Slightly greater gains may be seen for maximal muscle power, which typically demonstrates increases of 20-50% after months of HRST training both in young untrained persons [18,27,39], elite soccer players [4] (Fig.1) and old individuals [12,17]. Notably, HRST appears to evoke similar [17,18] or greater [4] gains in maximum muscle power than low-resistance types of resistance training (cf. Fig.1).

Fig.1 Peak quadriceps muscle power production before (full line) and after (dotted) 12 wks of heavy-resistance (8 RM, HRgroup) or low-resistance (24 RM, LRgroup) strength training in elite soccer players. Data from Aagaard et al. [4].
As a result of adaptive alterations in neural function [1,5,6], muscle morphology [2,21,29] and architecture [2] HRST is found to induce greater gains in maximal muscle strength and power compared to lowresistance types of training [3,4,18,27] (cf. Fig.1). Notably, the increase in maximum muscle strength contributes stiffness may increase with HRST [28,34], which theoretically allows to perform faster stretch-shortening actions (SSC) that for a given muscle work output would translate into an elevated power production.
Excised muscle fibers obtained from subjects exposed to years of resistance training demonstrate greater contractile force and power production compared to that measured in single muscle fibers from untrained age-matched individuals [37] (Fig.3). Maximum fiber shortening speed as well as muscle fiber power normalized relative to fiber size appear to remain unaffected by HRST [37,41], altogether suggesting that the superior in vitro power production of single muscle fibers with HRST is mainly the overall result of individual muscle fiber hypertrophy.
Eccentric training
Neuromuscular activity is suppressed during maximal eccentric (ECC) muscle contraction in untrained subjects [5,36], due to reduced levels of central activation [9] and reduced efferent motorneuron outflow indicated by diminished evoked V wave responses [19]. Maximum ECC muscle force during MVC can be increased by superimposed electrical muscle stimulation only in untrained individuals and not in trained strength athletes, indicating that the suppression in motorneuron activation may be altered by strength training [7]. In support of this notion, maximum ECC muscle strength was increased by use of HRST [3,13] due to a removed or diminished suppression in neuromuscular activity [5,8]. Notably, HRST was found to produce elevated H-reflex and V wave responses during MVC, indicating increased excitability of spinal motorneurons, decreased presynaptic inhibition and elevated descending motor drive [6,16]. Notably, the use of supramaximal ECC strength training leads to selectively elevated V wave responses during maximal eccentric contraction [19], demonstrating that adaptive changes in spinal motorneuron function and/or descending motor drive were evoked during ECC contraction. As a result, maximal ECC muscle strength is markedly increased with this particular type of training (discussed below).
Eccentric resistance training using submaximal, maximal (100% 1RM) or supramaximal (> 100% 1RM) training loads may lead to greater increases in maximal muscle strength compared to more conventional types of HRST [13,23,24,25,33,35], in part due to greater or a more pronounced degree of muscle hypertrophy following eccentric training [23-25,33] (Fig.2).

Fig.2 Changes in quadriceps muscle cross-sectional area after conventional (grey bars) and supramaximal eccentric (black bars) strength training. Data from Norrbrand et al. [33].
The greater gains in maximal muscle strength is expected to result in greater increases in maximal muscle power, since power is constituted by the product between instantaneous muscle force and muscle shortening speed, respectively. Animal experiments have shown that eccentric resistance training, especially when performed at long fiber lengths (i.e. at highly flexed joint positions, such as in deep squats) may lead to sarcomere addition and elongated muscle fiber lengths [10,11,30,31]. Notably, elongated muscle fiber lengths will contribute substantially to the gain in maximal muscle power induced by training. Thus, simulation analysis using empirical F-V and P-V relationships obtained for single human muscle fibers reveals that a 10% increase in maximal isometric muscle fiber force leads to a 10% gain in maximum (peak) power production, while a 10% increase in fiber length with a corresponding increase in maximal fiber shortening speed (Vo) cause maximum (peak) power to also increase by 10%. However, the relative gain in muscle power generated at high contraction speed (corresponding to 75% of Vo) is 4-fold higher (42% vs 10%) when comparing the effect of increasing muscle fiber length by 10% vs increasing maximum isometric force by 10%, respectively. Most potently, however, if a 10% gain is achieved both for maximum force and muscle fiber length, respectively, this yields a 56% gain in high-speed power (at 75% Vo). In comparison, maximum (peak) power production is increased by 21%. Thus, any training induced increase in muscle fiber length is likely to have a strong positive influence on the magnitude of maximal muscle power production, especially during very fast movements.
Plyometric training
Plyometric training involves forceful stretch-shortening (SSC) actions in the muscle-tendon complex [26] that typically involves production of high eccentric muscle forces, and where the magnitude of peak muscle loading (net joint moments) is remarkably similar to those observed during conventional HRST exercises [38]. Consequently, plyometric training may lead to muscle hypertrophy, at least when performed in previously untrained subjects [32,40], while frequently also leading to more pronounced gains in maximal muscle power than that attained by HRST alone [15]. However, the latter finding may be obscured by the fact that plyometric training typically introduces a large learning effect when subjects are evaluated (tested) in SSC exercises that are identical to those used in the plyometric training. Regardless, for athletes involved in explosive-type events such as sprinting, maximal jumping and throwing, the combination of HRST and plyometric training may lead to enhanced performance and greater gains in maximal muscle power than HRST alone.
Single muscle fiber data obtained in previously untrained subjects (physical education students) before and after plyometric training suggest this training modality results in an elevated maximum muscle fiber shortening speed in vitro, which explains the substantial gain in maximal power production observed for isolated muscle fibers [32].
The optimal loading intensity for maximizing the gain in muscle power with plyometric training remains a subject of debate [15]. In addition, the optimum power load (i.e. the load eliciting maximal power output) varies between muscle groups, and is also affected by the training status of the subject, as well as the choice of exercise [15]. For example, in strength trained individuals maximum leg muscle power was found to occur during unloaded (0% 1RM) squat jumping whereas the optimum load for maximizing the power output during conventional squat exercise was 40-70% of 1RM [14]. Nevertheless, the optimal training load for inducing maximal longitudinal gains in muscle power may well include very high loading intensities of 80-100% of 1RM (cf. Fig.1) to ensure that optimal adaptation take place both in neural function [6,22] and muscle size [20], and while performed in combination with eccentric training and/or plyometric training exercises. The functional significance of the latter training modality may primarily be to evoke increases in muscle fiber length, and to train ('learn') the CNS to perform very rapid SSC movements.

Fig.3 Force-velocity and power-velocity relationships (group mean curves) obtained for single isolated muscle fibers from long-term resistance trained (RT; 7.6 +/- 1.6 yrs of RT, full lines) and age matched untrained (NT, dotted lines)
individuals. Data from Shoepe et al. [37].
Conclusions
Maximal muscle strength and power are elevated in response to heavy-resistance strength training (HRST), and the addition of eccentric and/or plyometric training appear to evoke enhanced gains in these parameters. The adaptive physiological mechanisms involve changes in neuromuscular activity and alterations in muscle morphology (fiber size, fiber lengths) and muscle architecture, respectively, while changes in tendon stiffness may potentially contribute as well.
References:
[1] Aagaard P. Exerc. Sports Sci. Reviews 31, 61-67, 2003
[2] Aagaard P, Andersen JL, Dyhre-Poulsen P, Leffers AM, Wagner A, Magnusson SP, Halkjaer-Kristensen J, Simonsen EB. J. Physiol. 534.2, 613-623, 2001
[3] Aagaard P, Simonsen EB, Trolle M, Bangsbo J, Klausen K. Acta Physiol. Scand. 156, 123-129, 1996
[4] Aagaard P, Simonsen EB, Trolle M, Bangsbo J, Klausen K. Eur. J. Appl. Physiol. 69, 382-386, 1994
[5] Aagaard P, Simonsen EB, Andersen JL, Magnusson P, Halkjær-Kristensen J, Dyhre-Poulsen P. Appl. Physiol. 89, 2249-2257, 2000
[6] Aagaard P, Simonsen EB, Andersen JL, Magnusson SP, Dyhre-Poulsen P. J. Appl. Physiol. 92, 2309-2318, 2002
[7] Amiridis IG, Martin A, Morlon B, Martin L, Cometti G, Pousson M, van Hoecke J. Eur. J. Appl. Physiol. 73, 149-56, 1996
[8] Andersen LL, Andersen JL, Magnusson SP, Aagaard P. Eur. J. Appl. Physiol. 93, 511-518, 2005
[9] Babault N, Pousson M, Ballay Y, Van Hoecke J. J. Appl. Physiol. 91, 2628–2634, 2001
[10] Butterfield TA, Leonard TR, Herzog W. J. Appl. Physiol. 99, 1352-1358, 2005
[11] Butterfield TA, Herzog W. Pflugers Arch. 451, 688-700, 2006
[12] Caserotti P, Aagaard P, Larsen JB, Puggaard P. Scand. J. Med. Sci. Sports in press, 2008
[13] Colliander EB, Tesch PA. Acta Physiol. Scand. 140, 31-39, 1990
[14] Cormie P, McCaulley GO, Triplett NT, McBride JM. Med. Sci. Sports Exerc. 39, 340-349, 2007
[15] Cronin J, Sleivert G. Sports Med. 35, 213-234, 2005
[16] Del Balso C, Cafarelli E. Appl. Physiol. 103, 402–411, 2007
[17] de Vos NJ, Singh NA, Ross DA, Stavrinos TM, Orr R, Fiatarone Singh MA. J. Gerontol. 60A, 638–647, 2005
[18] Duchateau J, Hainaut K. J. Appl. Physiol. 56, 296-301, 1984
[19] Duclay J, Martin A, Robbe A, Pousson M. Med. Sci. Sports Exerc. 40, 722–734, 2008
[20] Fry AC. Sports Med. 34, 663-679, 2004
[21] Häkkinen K, Newton RU, Gordon SE, McCormick M, Volek JS, Nindl BC, Gotshalk LA, Campbell WW, Evans WJ, Häkkinen A, Humphries BJ, Kraemer WJ. J. Gerontol. 53, B415-423, 1998
[22] Häkkinen K, Komi OV, Alén M, Kauhanen H. Eur. J. Appl. Physiol. 56, 419-427, 1987
[23] Higbie EJ, Cureton KJ, Warren GL, Prior BM. J. Appl. Physiol. 81: 2173-2181, 1996
[24] Hortobagyi T, Hill JP, Houmard JA, Fraser DD, Lambert NJ, Israel RG. J. Appl. Physiol. 80, 765-772, 1996
[25] Hortobagyi T, Dempsey L, Fraser D, Zheng D, Hamilton G, Lambert J, Dohm L. J. Physiol. 524.1, 293-304, 2000
[26] Ishikawa M, Finni T, Komi PV. Acta Physiol Scand. 178, 205-213, 2003
[27] Kaneko M, Fuchimoto T, Toji H, et al. Scand. J. Sports Sci. 5, 50-55, 1983
[28] Kongsgaard M, Reitelseder S, Pedersen TG, Holm L, Aagaard P, Kjaer M, Magnusson SP. Acta Physiol. [Oxf.] 191, 111-121, 2007
[29] Kraemer WJ, Patton JF, Gordon SE, Harman EA, Deschenes MR, Reynolds K, Newton RU, Triplett NT, Dziados JE. J. Appl. Physiol. 78, 976-989, 1995
[30] Lynn R, Morgan DL. J. Appl. Physiol. 77, 1439-1444, 1994
[31] Lynn R, Talbot JA, Morgan DL. J. Appl. Physiol. 85, 98-104, 1998
[32] Malisoux L, Francaux M, Nielens H, Theisen D. J. Appl. Physiol. 100, 771-779, 2006
[33] Norrbrand L, Fluckey JD, Pozzo M, Tesch PA. Eur. J. Appl. Physiol. 102, 271–281, 2008
[34] Reeves ND, Maganaris CN, Narici MV. J. Physiol. 548, 971–981, 2003
[35] Seger JY, Arvidson B, Thorstensson A. Eur. J. Appl. Physiol. 79, 49-57, 1998
[36] Seger JY, Thorstensson A. Eur. J. Appl. Physiol. 69, 81-87, 1994
[37] Shoepe TC, Stelzer JE, Garner DP, Widrick JJ. Med. Sci. Sports Exerc. 35, 944–951, 2003
[38] Sørensen H, Lønbro S, Brink M, Vissing K, Overgaard K, Aagaard P. Proceedings 6th International Conference on Strength Training [ICST 2008], Colorado Springs, USA
[39] Toji H, Suei K, Kaneko M. Can. J. Appl. Physiol. 22, 328-336, 1997
[40] Vissing K, Brink M, Lønbro S, Sørensen H, Overgaard K, Danborg K, Mortensen J, Elstrøm O, Rosenhøj N, Ringgaard S, Andersen J, Aagaard P. J. Strength Cond. Res. 22, 1799–1810, 2008
[41] Widrick, Jeffrey J., Julian E. Stelzer, Todd C. Shoepe, and Dena P. Garner. Am. J. Physiol. 283, R408–R416, 2002

Valg og periodisering af repetitionstempo med henblik på optimal stimulering af muskeltilvækst.
Skrevet 17. aug 2012, 14:20